Now Reading: G-CSF Aracılığıyla Omentumda Metastatik Niş Gelişimi
-
01
G-CSF Aracılığıyla Omentumda Metastatik Niş Gelişimi
Omentum, karın boşluğu içindeki organlardan kaynaklanan birçok kanser türü için, özellikle de epitelyal yumurtalık, gastrik, kolorektal ve pankreas kanserleri için yaygın bir metastatik yayılım bölgesidir [1]. Bu belirgin tropizmin altında yatan nedenler, basit bir pasif hücre birikiminin ötesinde, anatomik özellikler, hücresel adezyon molekülleri, lokal mikroçevresel faktörler ve bağışıklık sistemi etkileşimlerini içeren karmaşık ve çok faktörlü mekanizmalar bütünüdür. Omental metastazın başarılı bir şekilde gerçekleşmesi, kanser hücrelerinin primer tümörden ayrılması, periton boşluğunda hayatta kalması, omentuma yönelmesi, tutunması ve nihayetinde invazyon yaparak çoğalmasını gerektiren çok adımlı bir kaskadı içerir.
Omentumun anatomik yapısı ve fizyolojik özellikleri, metastatik hücrelerin buraya yönelmesinde ve tutunmasında başlangıç rolü oynar. Periton sıvısının karın boşluğu içindeki doğal sirkülasyonu, askıdaki kanser hücrelerini yerçekimi ve diyafram hareketleri etkisiyle omentumun yüzeyine doğru yönlendirebilir. Omentumun geniş yüzey alanı ve lenfatik drenajdaki rolü, sıvı ve hücrelerin burada birikmesine katkıda bulunur. Ayrıca, omentumun yüzeyinde bulunan ve “milky spots” (süt lekeleri) olarak adlandırılan, bağışıklık hücreleri ve zengin kan damarı ağları içeren özelleşmiş yapılar, kanser hücrelerinin ilk tutunma ve yerleşme noktaları olarak işlev görür [2, 3]. Bu bölgelerdeki yüksek endotelyal venüller (HEV’ler) gibi damarsal yapılar, hem bağışıklık hücrelerinin hem de potansiyel olarak kanser hücrelerinin dolaşımdan dokuya geçişini kolaylaştırabilir [1].
Kanser hücrelerinin omentum yüzeyine fiziksel olarak tutunması, metastatik sürecin kritik bir adımıdır ve spesifik moleküler etkileşimler aracılığıyla gerçekleşir. Kanser hücrelerinin yüzeyinde eksprese edilen CD44 gibi adezyon molekülleri, omentumu kaplayan mezotelyal hücrelerdeki hyaluronan gibi ligandlara bağlanarak ilk teması sağlar [1]. Benzer şekilde, integrinler (örneğin α5β1) gibi diğer adezyon reseptörleri, mezotelyal hücreler arasındaki veya altındaki ekstraselüler matriks (ECM) bileşenlerine (örneğin fibronectin) bağlanarak tutunmayı güçlendirir [1]. Bazı durumlarda, kanser hücreleri mezotelyal hücre tabakasının bütünlüğünü bozarak veya mezotelyal hücrelerin dökülmesini tetikleyerek altındaki ECM’ye doğrudan erişim sağlayabilir ve invazyonu başlatabilir.
Omentumun mikroçevresi, sadece pasif bir tutunma yüzeyi olmaktan öte, metastatik hücrelerin hayatta kalması, çoğalması ve invazyonu için aktif olarak destekleyici bir ortam sunar. Omentumun ana bileşeni olan adipositler (yağ hücreleri), kanser hücreleri için önemli bir enerji kaynağı olan lipidleri sağlar ve adipokinler gibi çeşitli sinyal molekülleri salgılayarak tümör büyümesini teşvik eder [1]. Omental fibroblastlar ve mezenkimal kök hücreler gibi diğer stromal hücreler de büyüme faktörleri (örneğin VEGF) ve sitokinler salgılayarak anjiyogenezi (yeni damar oluşumu) uyarır ve kanser hücrelerinin proliferasyonunu destekler [1]. Bu stromal etkileşimler, metastatik lezyonların hızla büyümesi için kritik öneme sahiptir.
Omental mikroçevredeki bağışıklık hücreleri de metastaz sürecinde çift yönlü roller oynayabilir. Özellikle tümör ilişkili makrofajlar (TAM’lar), omentumda bol miktarda bulunur ve metastatik nişin şekillenmesinde önemli bir oyuncudur [1]. Başlangıçta M1 benzeri anti-tümör aktivite gösterebilseler de, sıklıkla M2 benzeri, pro-tümöral bir fenotipe dönüşürler. M2 makrofajları, anjiyogenezi teşvik eden, ECM’yi yeniden şekillendiren ve kanser hücrelerinin immün sistemden kaçışını kolaylaştıran faktörler salgılayarak metastatik büyümeyi destekler [1]. Dolayısıyla, omentumdaki immün manzara, metastatik hücrelerin yerleşmesi ve büyümesi için genellikle immün baskılayıcı ve destekleyici bir hale gelir.
Metastatik süreçte giderek daha fazla önem kazanan bir mekanizma, nötrofillerin rolü ve özellikle oluşturdukları Nötrofil Ekstraselüler Tuzakları (NET’ler) aracılığıyla metastatik nişi hazırlama ve kanser hücrelerini yakalama yetenekleridir [1]. Primer tümörden salgılanan çeşitli faktörler nötrofil akışını tetikler; bunlar arasında Granülosit Koloni Uyarıcı Faktör (G-CSF) özel bir yere sahiptir. G-CSF, bilinen temel görevi olan kemik iliğinden nötrofil üretimini ve salınımını artırmanın yanı sıra [4], kanser hücreleri tarafından da salgılanabilir ve güçlü bir kemoatraktan olarak nötrofillerin omentuma, özellikle milky spot bölgelerine hedeflenmiş göçünü uyarır [1]. Bu G-CSF kaynaklı nötrofil birikimi, metastatik hücreler gelmeden önce omentumda “premetastatik niş” oluşumunun kritik bir parçasıdır.
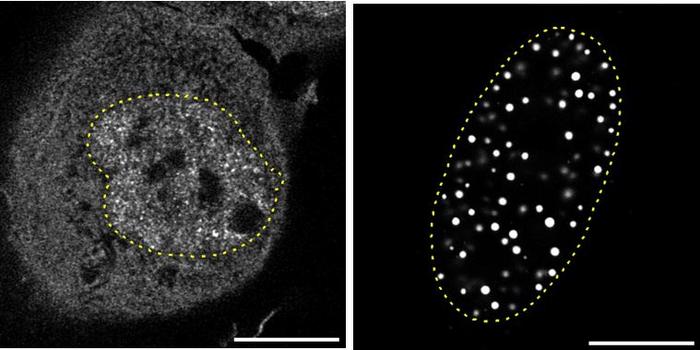
Omentuma yönlendirilen ve biriken nötrofiller, yine primer tümörden kaynaklanan veya lokal olarak salınan faktörlerle aktive edilir. Özellikle G-CSF’nin, IL-8 ve GRO gibi diğer sitokinlerle birlikte, bu nötrofilleri NETosis adı verilen özelleşmiş hücre ölümü ve NET salınımı sürecine sokmada doğrudan etkili olduğu gösterilmiştir [1]. Bu durum, erken evre yumurtalık kanseri olan hastalarda ve henüz metastaz gelişmemiş tümörlü fare modellerinde, omentumda NET belirteçlerinin (örneğin sitrüline histon H3, hücre dışı DNA) varlığıyla doğrulanmıştır [1]. Bu, G-CSF dahil olmak üzere tümör kaynaklı faktörlerin sadece nötrofilleri bölgeye çekmekle kalmayıp, aynı zamanda onları metastazı kolaylaştırıcı NET yapılarını oluşturmaları için programladığını gösterir.
NETosis’in moleküler mekanizması, özellikle G-CSF gibi faktörlerin etkisi altında daha da belirginleşir. G-CSF’nin ve benzer şekilde IL-8 ile GRO’ların nötrofilleri uyarması, hücre içi sinyal yollarını aktive ederek ‘respiratuar burst’ adı verilen olayı güçlü bir şekilde potansiyelize eder ve bu da reaktif oksijen türlerinin (ROS) bol miktarda üretimine yol açar [6, 7]. G-CSF’nin bu etkisi, tek başına veya diğer sitokinlerle sinerjistik olarak çalışarak, NET oluşumu için kritik bir eşik olan ROS seviyelerine ulaşılmasını kolaylaştırır. Artan ROS seviyeleri, çekirdekteki Peptidilarginin Deiminaz 4 (PAD4) enziminin aktivasyonunu tetikler [8]. PAD4 enzimi, histon H3 gibi proteinlerdeki argininleri sitrüline çevirir [9]. Bu sitrülinasyon, kromatin yapısının çözülmesine (dekondensasyon) neden olur ve sonuçta DNA iplikçikleri, modifiye histonlar ve granül proteinleri hücre dışına atılarak NET ağlarını meydana getirir [5, 9]. Bu sürecin spesifikliği, MCP-1 gibi bazı kemokinlerin nötrofil göçünü uyarırken NETosis’i belirgin şekilde tetiklememesiyle ortaya konmuştur [1, 10].
Omentumda oluşan NET’lerin metastazdaki etkisi özellikle çarpıcıdır. Bu yapışkan, ağ benzeri yapılar, periton sıvısında dolaşan kanser hücreleri için son derece etkili fiziksel tuzaklar olarak işlev görür [1, 5]. NET’lerin DNA iskeleti ve üzerine yapışmış proteinler, hareket halindeki kanser hücrelerini mekanik olarak yakalar ve omentum yüzeyine sabitler. Bu yakalama mekanizması, kanser hücrelerinin akışkan ortamdan durağan bir yüzeye geçiş yapabilmesi ve kolonizasyon sürecini başlatabilmesi için kritik bir öneme sahiptir. Kanser hücrelerinin bu NET ağlarına bağlandığı ve bu etkileşimin hücre tutunmasını artırdığı deneysel olarak gösterilmiştir [1]. Bu fiziksel engelleme ve tutunma sağlama işlevi, NET’leri omental metastaz kaskadında merkezi bir oyuncu haline getirir.
NET’lerin metastazı destekleyici etkisi, sadece mekanik yakalama ile sınırlı değildir. NET’ler üzerinde yoğunlaşmış olan nötrofil elastaz ve MMP gibi proteolitik enzimler, çevrelerindeki ekstraselüler matriksi parçalayabilir ve yeniden şekillendirebilir [11]. Bu durum, yakalanan kanser hücrelerinin omentum dokusunun daha derinlerine doğru invazyon yapmasını kolaylaştırabilir. Ayrıca, NET bileşenleri veya NET oluşumu sırasında salınan diğer moleküller, lokal inflamatuar ortamı daha da pro-tümöral hale getirebilir veya dormant (uykuda) kanser hücrelerini uyandırarak metastatik büyümeyi tetikleyebilir [11]. NET yapılarının ortadan kaldırılmasının (PAD4 genetik eksikliği veya PAD4 inhibitörleri/DNaz kullanımı ile) omental metastazı dramatik bir şekilde azaltması, bu yapıların metastatik süreçteki nedensel ve kritik rolünü güçlü bir şekilde kanıtlamaktadır [1].
Omental metastazın altında yatan mekanizmaların çeşitliliği, bu sürecin karmaşıklığını ve hedefe yönelik tedavi geliştirmenin zorluğunu ortaya koymaktadır. Anatomik faktörler, hücre adezyon molekülleri, stromal hücre etkileşimleri, makrofajlar gibi immün hücreler ve özellikle G-CSF gibi faktörlerle aktive edilen ve NET oluşturan nötrofiller, bu sürecin farklı aşamalarında rol oynayan önemli bileşenlerdir. Farklı kanser türlerinde bu mekanizmaların göreceli önemleri değişkenlik gösterebilir. Bu nedenle, metastazı engellemeye yönelik terapötik stratejiler geliştirilirken, bu çok yönlü mekanizmaların tümü göz önünde bulundurulmalıdır.
Özellikle, kemoterapiye bağlı nötropeninin tedavisinde standart olarak kullanılan G-CSF’nin çift taraflı rolü dikkate alınmalıdır. Bir yandan nötropeniyi düzelterek enfeksiyon riskini azaltırken, diğer yandan nötrofilleri aktive edip NET oluşumunu tetikleyerek omental metastaz potansiyelini artırabilir [1]. Hatta bazı klinik çalışmalarda, profilaktik G-CSF kullanımının yumurtalık kanseri hastalarında sağkalım avantajı sağlamadığı gözlemlenmiştir [1]. Bu durum, G-CSF tedavisinin özellikle metastaz riski taşıyan hastalarda dikkatli bir risk-fayda değerlendirmesiyle uygulanması gerektiğini vurgular. NET oluşumunu hedefleyen tedaviler veya G-CSF’nin metastazı teşvik edici etkilerini önleyecek yaklaşımlar, gelecekteki kanser tedavileri için önemli araştırma alanlarıdır.
Referanslar
1. Lee, W., Ko, S. Y., Mohamed, M. S., Kenny, H. A., Lengyel, E., & Naora, H. (2018). Neutrophils facilitate ovarian cancer premetastatic niche formation in the omentum. Journal of Experimental Medicine, 216(1), 176–194. https://doi.org/10.1084/jem.20181170
2. Hagiwara, A., Takahashi, T., Sawai, K., Taniguchi, H., Shimotsuma, M., Okano, S., Sakakura, C., Tsujimoto, H., Osaki, K., & Sasaki, S. (1993). Milky spots as the implantation site for malignant cells in peritoneal dissemination in mice. Cancer Research, 53(3), 687–692.
3. Meza-Perez, S., & Randall, T. D. (2017). Immunological functions of the omentum. Trends in Immunology, 38(7), 526–536. https://doi.org/10.1016/j.it.2017.03.002
4. Negus, R. P., Stamp, G. W., Relf, M. G., Burke, F., Malik, S. T., Bernasconi, S., Allavena, P., Sozzani, S., Mantovani, A., & Balkwill, F. R. (1995). The detection and localization of monocyte chemoattractant protein-1 (MCP-1) in human ovarian cancer. Journal of Clinical Investigation, 95(5), 2391–2396. https://doi.org/10.1172/JCI117933
5. Brinkmann, V., Reichard, U., Goosmann, C., Fauler, B., Uhlemann, Y., Weiss, D. S., Weinrauch, Y., & Zychlinsky, A. (2004). Neutrophil extracellular traps kill bacteria. Science, 303(5663), 1532–1535. https://doi.org/10.1126/science.1092385
6. Fuchs, T. A., Abed, U., Goosmann, C., Hurwitz, R., Schulze, I., Wahn, V., Weinrauch, Y., Brinkmann, V., & Zychlinsky, A. (2007). Novel cell death program leads to neutrophil extracellular traps. Journal of Cell Biology, 176(2), 231–241. https://doi.org/10.1083/jcb.200606027
7. Nathan, C. F. (1989). Respiratory burst in adherent human neutrophils: Triggering by colony-stimulating factors CSF-GM and CSF-G. Blood, 73(1), 301–306.
8. Li, P., Li, M., Lindberg, M. R., Kennett, M. J., Xiong, N., & Wang, Y. (2010). PAD4 is essential for antibacterial innate immunity mediated by neutrophil extracellular traps. Journal of Experimental Medicine, 207(9), 1853–1862. https://doi.org/10.1084/jem.20100239
9. Wang, Y., Li, M., Stadler, S., Correll, S., Li, P., Wang, D., Hayama, R., Leonelli, L., Han, H., & Grigoryev, S. A. (2009). Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. Journal of Cell Biology, 184(2), 205–213. https://doi.org/10.1083/jcb.200806072
10. Rollins, B. J., Walz, A., & Baggiolini, M. (1991). Recombinant human MCP-1/JE induces chemotaxis, calcium flux, and the respiratory burst in human monocytes. Blood, 78(4), 1112–1116.
11. Albrengues, J., Shields, M. A., Ng, D., Park, C. G., Ambrico, A., Poindexter, M. E., Upadhyay, P., Uyeminami, D. L., Pommier, A., & Küttner, V. (2018). Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science, 361(6408), eaao4227. https://doi.org/10.1126/science.aao4227